Электрофорез в агарозном геле проводят в горизонтальном направлении, так как при этом 1) гель с низкой концентрацией агарозы лучше держится, 2) получается меньшее перекашивание (коллапс) в процессе электрофореза и 3) меньше искажаются полосы ДНК. По-видимому, проще всего работать с системой, когда гель полностью покрыт слоем буфера для электрофореза толщиной около 1 мм. Сопротивление агарозы ненамного превышает сопротивление буфера, так что значительная часть тока течет через агарозу.
Во все буферы с нейтральными значениями рН можно добавлять 0,5 мкг/мл бромистого этидия.
В процессе электрофореза буфер у анода защелачивается, а у катода закисляется. Поэтому обычно пользуются буферной системой с высокой емкостью. Буферы не содержат ионов Cl — , так как последние не обладают буферной емкостью, и их присутствие может привести к тому, что ДНК в геле утратит биологическую активность.
- Трис-боратный буфер. Обладает высокой буферной емкостью. При его использовании, по-видимому, получаются наиболее узкие полосы. Основной раствор 10 × не зарастает, но при длительном хранении в нем образуется осадок, который растворим в щелочи. В присутствии бората агарозные гели не растворяются при высокой концентрации NaClO4 или KI.
- Трис-фосфатный буфер. Тоже обладает высокой емкостью. При его использовании получаются примерно такие же результаты, что и при использовании трис-боратного буфера. Основной раствор, однако, может зарастать. Его преимущество заключается в том, что такие гели можно растворять в концентрированном растворе NaClO4 или KI.
- Трис-ацетатный буфер. Обладает относительно низкой буферной емкостью, и при длительном электрофорезе может возникнуть необходимость в рециркуляции буфера в аппарате. Использование при электрофорезе высоких напряжений не вызывает нагрева. Исходные растворы могут зарастать. Гели в трис-ацетатном буфере можно растворять в концентрированном NaClO4 или KI. Этот буфер, по-видимому, используется наиболее широко.
- Щелочной буфер. Обладает очень низкой емкостью. Обычно при его использовании необходима рециркуляция буфера в аппарате. Наносимые образцы не должны содержать Mg 2+ , иначе ДНК выпадет в осадок. Ионы Mg 2+ необходимо удалять, добавляя избыточные количества ЭДТА.
Препараты агарозы значительно различаются по твердости, по разрешающей способности при разделении фрагментов ДНК, электрофоретической подвижности ДНК, легкости плавления, прозрачности и наличию вредных примесей. Наиболее вредной примесью являются, по-видимому, сульфонированные агарозы, подавляющие активность многих ферментов, работающих на нуклеиновых кислотах.
Обычно мы пользуемся агарозой фирмы МС/В. Агарозу растворяют в буфере для электрофореза, нагревая до кипения в микроволновой печи. Необходимо убедиться, что раствор стал гомогенным и что в нем не осталось твердых частиц агарозы. Перед тем как залить гель, раствор охлаждают до 50°С. Если заливать гель слишком горячей агарозой, то легко можно повредить аппарат для электрофореза
Лунки для образцов делают с помощью погруженной в расплавленный гель гребенки из оргстекла, полихлорвинила или тефлона. Гребенку устанавливают до заливки геля таким образом, чтобы кончики зубьев находились примерно в 0,5 мм от основания геля. Если лунки достанут дна, то образец может протечь под гель. Когда гель полностью застынет, гребенку вынимают и лунки заполняют буфером для электрофореза.
Г. Нанесение образцов и краски
Наносимые образцы содержат 5-10% глицерола или 5-10% сахарозы и 0,025% красителя, благодаря которому можно проследить за ходом электрофореза. Например, можно добавить в образец 1 /10 объема раствора, содержащего 50% глицерола и 0,25% красителя.
ДНК. Наносите около 10 нг ДНК в расчете на каждую ожидаемую полосу. Гель будет перегружен, если в полосе окажется более чем примерно 100 нг ДНК.
Бромфеноловый синий (распадается в щелочи).
Бромкрезоловый зеленый (обладает одинаковой подвижностью и в нейтральных, и в щелочных растворах). Ксиленцианол FF (распадается в щелочи; подвижность ниже, чем подвижность бромфенолового или бромкрезолового).
Образцы можно наносить, используя автоматическую пипетку и полипропиленовый наконечник или с помощью микрошприца, на который надет тонкий пластиковый шланг.
Подвижность небольших молекул ДНК может быть такой же или даже большей, чем подвижность используемого красителя. Чем ниже концентрация агарозы и (или) выше напряженность, тем больший фрагмент ДНК будет обладать такой же подвижностью, как и краситель. Краситель поглощает флуоресценцию связанного с ДНК бромистого этидия. Это приводит к тому, что в том месте геля, где находится краска, невозможно наблюдать слабые полосы ДНК. Образцы можно наносить и без краски.
источник
Установка для электрофореза в агарозном геле.На рисунке 7 представлены основные элементы для проведения горизонтального электрофореза в агарозном геле. Схематично, установку можно представить как емкость для электродного буфера, по краям которой расположены два электрода. В пространство между электродами заливается проводящий буфер, в котором проводится электрофорез, и помещают гель. При этом гель может быть залит непосредственно в камере, а может быть залит отдельно на специальном лотке, который помещается в камеру.
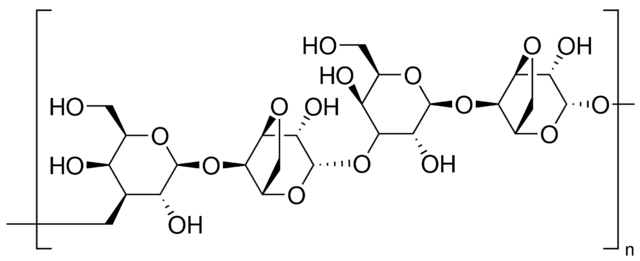
Агароза. Природный коллоид, который выделяют из морских водорослей, является линейным полисахаридом, образованным повторяющимся элементом ‒ агаробиозой, которая в свою очередь состоит из чередующихся элементов: галактозы и 3,6-ангидрогалактозы. Агароза очень хрупка, и легко разрушается при манипулировании. Агарозные гели имеют «поры» большого размера и используются преимущественно для разделения больших молекул.
Агарозные гели получают суспендированием сухого порошка агарозы в водном буфере, и кипячением смеси до того момента, когда агароза расплавится и образует прозрачный раствор. Затем раствор наливают на подложку и дают остыть до комнатной температуры, чтобы сформировался прочный гель. При застывании агароза формирует матрикс, плотность которого определяется концентрацией.
Буфер для электрофореза. На электрофоретическую подвижность ДНК воздействуют состав и ионная сила буфера для электрофореза. В отсутствии ионов, электропроводность минимальна, и перемещение ДНК происходит медленно. В буфере с высокой ионной силой электропроводность очень эффективна, и образуется значительное количество тепла. В худшем случае, гель расплавляется и ДНК денатурирует. Существует несколько буферов для электрофореза нативной двухцепочечной ДНК. Они содержат ЕОТА (рН 8,0) и Тгis-ацетат (ТАЕ), Тгis-борат (ТВЕ), или Тгis-фосфат (ТРЕ).
Маркерная ДНК. При заданном напряжении, концентрации агарозного геля и буфера, расстояние перемещения зависит от молекулярного веса исходного материала. Поэтому, маркерная ДНК известного размера должна наноситься на дорожки и с левого, и с правого края геля. Маркер обычно содержит определенный набор известных сегментов ДНК, которые облегчают определение размера исследуемой ДНК, если какое-либо систематическое искривление геля возникнет во время электрофореза.
Буфер для нанесения. Образцы ДНК, которые будут наноситься на агарозный гель, сначала смешивают с буфером для нанесения, обычно содержащим воду, сахарозу и краситель (например ксиленцианол, бромфеноловый синий, бромкрезол зеленый и другие).
Окраска и фиксация
Для анализа результатов электрофореза, после его завершения, требуется, прежде всего, визуализовать картину распределения полос в геле. С этой целью могут применяться два основных подхода. Первый способ состоит в окрашивании ДНК специфическими красителями. Для этой цели существует большое разнообразие красителей, способных специфически окрашивать двунитевые или однонитевые участки ДНК, ДНК в составе различных комплексов или в определённой конформации. К достоинствам такого подхода можно отнести высокую чувствительность, избирательность окрашивания и дешевизну метода. Среди недостатков можно упомянуть относительную трудоёмкость процедуры, невозможность следить за ходом электрофореза в реальном времени, токсичность некоторых из широко применяемых красителей, воздействие красителей на структуру ДНК, и необходимую иногда предварительную фиксацию ДНК в геле.
Другой путь основан на способности самой ДНК поглощать свет в УФ диапазоне. Поэтому можно просканировать гель в УФ-свете и определить местоположение ДНК в геле. В настоящее время промышленностью выпускаются специальные комплексы, позволяющие следить за миграцией ДНК в геле в автоматическом режиме непосредственно в ходе электрофореза и получать оцифрованное изображение в любой момент времени. При использовании этого подхода предварительная фиксация ДНК в геле не требуется. К недостаткам подхода можно отнести малую избирательность метода и относительную дороговизну оборудования.
Наиболее распространенным красителем ДНК в экспериментах по электрофорезу является бромистый этидий. Этот краситель относится к соединениям, способным интеркалировать между парами оснований ДНК. Вещество является сильным канцерогеном и мутагеном, способным проникать через кожу. По этой причине работа с ним требует особой осторожности и соблюдения повышенных мер безопасности. При работе с бромистым этидием всегда необходимо использовать перчатки и маску. Отходы, содержащие бромистый этидий, требуют специальной утилизации.
ВЫПОЛНЕНИЕ РАБОТЫ
1. Для приготовления 1%-го геля ‒ смешайте 100 мл воды и 1 г агарозы и доведите раствор до кипения. Размешайте до ресуспендирования осевшей агарозы. В пластиковую кювету для геля установите гребенку под будущие лунки. Влейте агарозу в кювету. Агароза должна заполнить куветю полностью. Когда гель затвердеет, осторожно удалите гребенку.
2. Готовим камеру ‒ кладем пластиковую кювету с гелем в камеру, заливаем буфером так, чтобы гель скрылся под ним.
3. Вносим 5–10 мкл образца в лунки и закрываем верхней крышкой.
4. К черному штекеру присоединяем отрицательный полюс, а к красному положительный.
5. Включаем форез ‒ 120 В 20–30 мин. Возле электродов должны быть небольшие пузырьки, как индикатор, что форез пошел. ДНК двигается от минуса к плюсу, поэтому лунки с ДНК должны быть возле черного штекера (минуса).
6. Через 20–30 мин, выключаем прибор, открываем крышку и вытаскиваем кувету с гелем шипцапи или используем перчатки. В случае наличия в образце красителя, разделение можно будет увидеть невооруженным глазом, а при его отсутствии просматривают гель в УФ-свете на трансиллюминаторе и фотографируют.
Сделать вывод о проделанной работе. Зарисовать электрофореграмму.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Дайте понятие электрофорезу.
2. Дайте понятие электрофорезу в агарозном геле.
3. Физические принципы электрофореза в агарозном геле.
4. Перечислите параметры, от которых зависит движение заряженных частиц в агарозном геле.
5. Перечислите компоненты, необходимые для электрофореза в агарозном геле.
6. Основные элементы установки для электрофореза в агарозном геле.
7. Объясните методы анализа результатов электрофореза, их достоинства и недостатки.
8. К какому полюсу происходит движение молекул ДНК при электрофорезе в агарозном геле и почему?
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ — конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.
источник
Особенности работы с низкопроцентными (0.3%) гелями.
в плашку для электрофореза залить «подложку» — слой агарозы 1.5-2.0% толщиной
2-3 mm, дать застыть;
ВНИМАНИЕ! Работать в холодной комнате. Гель очень хрупкий, все манипуляции проводить только вместе с 2% подложкой; не лить буфер на поверхность геля. Низкопроцентные гели особенно чувствительны к перегрузке по количеству DNA на дорожку.
Разделение линейных молекул
Диапазон нормального разделения линейных dsDNA молекул для гелей с различной концентрацией агарозы:
|
Меньший предел определяется (в основном) диффузией полосы в геле. Т.е. в гелях с низкой концентрацией агарозы мелкие фрагменты вполне разделяются, но полосы не четкие.
Верхний предел сильно зависит от напряженности поля, при которой проводится форез. Чем меньше напряженность поля, тем более длинные молекулы можно эффективно разделить.
Разделение суперскрученных и кольцевых молекул
К сожалению относительная подвижность линейных и кольцевых молекул зависит от условий фореза: % геля, скорость фореза (в частности, это означает, что нельзя пользоваться линейным маркером для оценки размера кольцевых молекул).
Приведенная таблица дает некоторое представление о соотношении подвижностей при умеренной (
6V/cm) скорости фореза (в скобках — при более быстром разгоне):
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В присутствии 0.5 µg/ml EtBr разрешение релаксированной и суперскрученной pDNA увеличивается примерно в 20 раз при повышении ионной силы буфера до 4хТАЕ. Того же увеличения можно добиться понижая концентрацию EtBr.
На 1% агарозном форезе ssDNA бежит чуть быстрее (
10%), чем dsDNA того же размера. ssDNA окрашивается EtBr заметно слабее, чем ds (разница в
4-5 раз). Tо есть, чтобы полосы имели одинаковую интенсивность окраски, требуется взять
Чтобы разделить цепи, нужно либо непосредственно перед форезом прогреть
1′ 100 o C, либо добавить к образцу NaOH до 0.1 М, 5-10′ (NT или 37 o C);
При оценке напряженности поля для горизонтального фореза принято пренебрегать конкретной геометрией камеры и измерять расстояние непосредственно между электродами.
На рисунке показана схема форезов, при разном напряжении. Форезы проводились разное время, так, чтобы 0.5kb фрагмент прошел одинаковое расстояние. Видно, что проведение фореза при высоком напряжении эквивалентно уменьшению длины геля.
Разумный компромисс между скоростью и качеством фореза для высококачественных или препаративных форезов:
2V/cm(можно меньше для ON форезов). Для аналитических форезов приемлемое качество сохраняется до
DNA особенно легко теряет EtBr при повышенной температуре (что обычно случается, если гонять форезы при высоком напряжении).
EtBr при форезе движется от (+) к (-). Если хочется, чтобы он не уходил из геля, лучше ввести его и в форезный буфер.
Количество DNA, которое можно наносить на дорожку
Нижний предел определяется используемым методом детекции. Если применяется окрашивание EtBr, то не стоит надеяться увидеть 2 O; рабочая концентрация в 1х буфере
0.005-0.02%) могут заметно мешать наблюдению фрагментов под UV. По нашему опыту, элюция фрагмента в РEG вместе с любым из этих красителей не оказывает заметного влияния на меченье, лигирование и трансформацию.
Cresol red (Na соль) — (стоковый раствор 50mM в H 2 O; рабочая концентрация в 1х буфере
0.1mM) совместим с ферментативными реакциями, практически не мешает наблюдению под UV.
OrangeG — (стоковый раствор 1% в формамиде; рабочая концентрация в 1х буфере
0.01-0.05%) наиболее подвижный краситель, практически всегда находится вне «рабочей зоны». Заметен под UV.
Краситель в буфере нужен лишь для того, чтобы образец был легко заметен в лунке и в геле. Обычно приводимые в методиках
0.025% для бромфенолового синего и ксиленцианола (в 1х буфере) по нашему мнению слишком большое количество.
* буфера с различными красителями.
* различные разведения буфера для внесения (10х, 2х, 1х). При этом образец любого объема собирается из двух компонентов (буфер+образец), а не из трех (буфер+ вода + образец).
1x если объем образца 25-75%;
10x ==> 75%
Смотри также:
/ссылки на сетевые ресурсы/
| Дополнения, комментарии, вопросы Для оптимального разделения не очень больших фрагментов, можно пользоваться буфером для внесения по рецепту, указанному в каталоге MBI Fermentas: 6x Orange Loading Dye Solution (#R0631) При этом ксиленцианол мигрирует с > 4 kb, а оранжевый — с 50 bp. Такой подход позволяет оптимизировать рабочее напряжение, а, соответственно, и разделение. И еще — я человек в мол.биологии непросвещенный, а потому да простят меня профессионалы, если я скажу всем известную вещь: вносить образцы в лунки гораздо проще, если под камеру подложить листок темной (черной или синей) бумаги, чтобы оттеняла лунки. Люди, хелп! Если кто-нибудь знает что-то про технологию внесения биополимеров в гель под названием Shark Teeth, напишите, пожалуйста! А почему бы заодно не добавить в текст, что рабочая концентрация бромистого этидия, например, 0,5 мкг/мл и что его можно добавлять прямо в гель, что гель с добавленным в него EthBr спокойно стоит ночь под электродным буфером без заметного вымывания из него краски? Полезные, вобшем-то факты, хоть и мелкие, но, с моей точки зрения, вполне не лишние. Акульи зубы — не гель. а гребенка в виде этих самых зубок. Втыкается в гель, и не вынимается, а образец наносится тонким-тонким носом в дырки между зубами. См. инструкцию к Макрофору. Только это не про агарозный форез. 2Re: что это гребенка такая, я знаю. Только сейчас вижу, что по формулировке моего вопроса кажется, будто это гель так называется:) Все, спасибо, уже поздно:) Только по моим данным, это имено про форез. Не знаю уж, про агарозный или ПААГ, по-моему, в данном случае не суть важно Народ, а какова длина волны для визуализации ПЦР-фрагментов? народ, а как точно называется маркер мол веса (лестница)? и куда, как ее наносить? а почему в гел Этьбромид а не GelStar ? GelGreen ? GelRed ? SYBRSafe ? а почему ТБЕ ТАЕ а не SB (sodium borate) ? а почему UV а не Blue ( Safe Imager , Dark Reader , FlashGel) А ПОКОЧАНУ )))) ответ от радио Molbiol — Не выпендрюйся Василий Иваныч-слушай свои «Валенки» 1: Biotechniques. 2004 Feb;36(2):214-6. Links Erratum in: Johns Hopkins University School of Medicine, Baltimore, MD, USA. PMID: 14989083 [PubMed — indexed for MEDLINE] All: 1 1: Electrophoresis. 2001 Mar;22(5):814-28.Click here to read Links Clare Chemical Research, Inc, Denver, CO 80206, USA. mseville@clarechemical.com The Dark Reader optical system (Clare Chemical Research, Denver, CO, USA) uses relatively low intensity broad-band visible blue light in combination with broad-band optical filters to detect fluorescence with a level of sensitivity that often surpasses that of UV transilluminators and can rival that of laser-based scanners. Applications of DR (Clare Chemical Research) devices include the detection of DNA and SYBR-stained protein samples following, and also during, electrophoresis. Unlike laser-based imaging systems, the fluorescence is directly visible to the user as well as being fully compatible with charge-coupled device (CCD) and Polaroid camera-based detection and imaging. Additionally, the DR optical system functions well in multicolor fluorophor environments. Because the Dark Reader does not emit any UV light, the extent of DNA damage incurred when visualizing DNA samples is drastically reduced compared to the damage produced by a UV device and this can have a significant benefit on downstream cloning protocols. Furthermore, dye photobleaching is minimal, extending the length of time that a fluorescent sample is visible. The inherent flexibility of the DR optical system allows many different configurations of the Dark Reader to be constructed such as transilluminators, hand lamps and integrated transilluminator-electrophoresis units. PMID: 11332748 [PubMed — indexed for MEDLINE] Today’s Featured Review Until recently Sybr green had been our labs nucleic stain of choice. We have, however, recently migrated to Biotium’s GelRed. GelRed is a newer nucleic acid gel stain that is safer and more temperature stable than other dyes. GelRed is safer because it is non mutagenic and is diluted in water. It is more thermostable than Sybr dyes because it does not degrade with repeated freeze-thaws or long storage (couple months at RT) and can be stored at room temperature, and sensitivity seems to at least equal if not exceed Sybr green when visualized with UV transilluminator. The dye is used similarly to Sybr dyes with staining (available at 10,000X ) or direct sample additions. The technology is not particularly ground breaking or new but the safety profile of this product makes it a viable alternative to potentially more dangerous dyes. Review by Vitelli Using GelGreen ‘In-Gel’ Save even more Time. GelGreen Safety FlashGel™ DNA System The FlashGel System is the fastest way to separate DNA and the only way to watch DNA migration as it happens. This revolutionary new tool separates DNA in 2 – 7 minutes. Monitor gel runs right at the bench, without UV light. The highly sensitive system allows a 5X reduction in DNA levels – saving both time and money. * Get results in 5 minutes or less. The FlashGel System consists of enclosed, disposable, precast agarose gel cassettes and a combination electrophoresis and transilluminator unit. * FlashGel Cassettes contain precast, prestained agarose gels and buffer – no need for gel preparation, buffer addition or gel staining. это все называется «ДНК технология» — не путайте с одноименной фирмой всем привет. Здравствуйте! «. При этом ксиленцианол мигрирует с > 4 kb, а оранжевый — с 50 bp. « а при какой процентности геля? Я так понимаю, как мигрируют красители именно от этого и зависит Как лучше выделить днк 200п.н. из геля для последующей амплификации. Скорее всего малая концентрация? Спасибо —Заранее спасибо ответившим, особенно ответившим правильно —. При этом ксиленцианол мигрирует с > 4 kb, а оранжевый — с 50 bp. « —а при какой процентности геля? Я так понимаю, как мигрируют красители именно от этого и зависит. Вроде красители в агарозе мигрируют на одно и тоже Rf не зависимо от процентности. А вот фрагменты сильно зависимо от процентности. Берете 2 маркера 100b и 1000b и в разных процентностях агарозы гоните с красителями и очень хорошо определяете, что с чем бежит. 3 процентный агароз вам уже трудно будет растворить. Я так дУмаю-у Мне кажется это лишняя работа элюировать ДНК из геля для последующей амплификации. Срез геля в воду, заморозить-разморозить и 1-2 мкл на ПЦР. Вы реамплифицируете кг. Спасибо, попробую на следующей неделе. А что если уже выделил из геля с помощью QG и при исходнике с приблизительной количеством 50 нг получил 40 мкг в 40 мкл. На форезе 1,2% в 10 мкл пробы ясен перец ничего не видно, агароза супер. Поставил на ПЦР, отправил 20 мкл в смесь, жду результатов. Сколько приблизительно может быть продукта? Спасибо Вопрос 1: Может ли в реакции сильнее амплифицироваться плазмида, чем нужная вставка?? 1% агарозу (в 2% линейная ДНК идет очень близко к суперскрученной, в 0.7% — близко к релаксированной). Если требуется проконтролировать циклизацию 3kbp фрагмента, то надо использовать 2% агарозу. если у Вас не нарабатывается большое кол-во продукта (20 нг в мкл и выше), дело уж конечно не в пирофосфатах- откуда бы им взяться, коли буквы не включились в амплификат (его мало)? Скорее всего проблемы с праймерами- посмотрите, нет ли «бомбы» (димер праймеров) внизу, или полимераза дохнет, что вря ли. Если проблема не лечится «в лоб», то просто замешайте исходно большой объем смеси, грубо говоря, накопите продукт экстенсивным путем, и всех делов. Я наверное перегрелся на солнцепёке (отпуск все-таки) — не понимаю в чем проблема плазмида или вставка, одна пара праймеров. По классике в ПЦР сильнее амплифицмруется всегда более короткий фрагмент. Без бутылки не разберешься, дождусь вечера. Здравствуйте, подскажите чем красить агарозный гель после щелочного электрофореза? И на какой стадии (до или после фиксирования в кислоте). Обязательно ли сушить? -Здравствуйте, подскажите чем красить агарозный гель после щелочного электрофореза? Бромий этидием или сибирским зеленым и иже с ними -до или после фиксирования в кислоте. «если я скажу всем известную вещь: вносить образцы в лунки гораздо проще, если под камеру подложить листок темной (черной или синей) бумаги, чтобы оттеняла лунки» Чем тогда думали в Сигма, если сделали столик в камере для фореза из белого пластика, к тому же скользкого в первом случае можно наклеить синей изоленты под место, где карманы)) Но от скольжения избавиться сложно — гель частенько уплывает от любых дуновений во время фореза. хоть гвоздями прибивай Можно ли заменить ТВЕ-буфер на Трис-СІ+EDTA для фореза ДНК, и какой молярности? За Трис.НСI в форезном буфере ставлю кол. Борная кислота испорчена у нас. Похоже, это ко мне . Поставляем наборы готовых гелей для электрофореза ДНК, рассчитанные на использование в ПЦРных лабораториях. В каждый комплект входит пузырёк ТАЭх20 (50 мл) для приготовления литра электрофоретического буферного раствора (EtBr 0,5 мкг/мл). Если нужно, то эти пузырёчки можно продавать и без гелей . Если интересно, то пишите. Ели реклама здесь неуместна, то удалите. Размер:: 19.19к кол-во скачиваний: 1724 Ну как может испортиться борная кислота? Купите тогда в аптеке. Борная кислота старая и «Ч.» по квалификации чистоты. Надо заказывать новые реактивы. Могу отсыпать сухой (белый порошок) ТВЕ. И дам пропись как самому готовить. Но про Трис.НСI для фореза — забудь. Лучше водопроводную воду. Забудьте и про Трис-HCl, и про все прочие буферы на основе Триса. Наиболее продвинутые пользователи за бугром переходят на Li-борат и Li-ацетат. Давно хотел попробовать карбонатный буфер, но всё руки не доходили. Недавно дошли. Приготовили Li-карбонатный буфер. Форезы получаются отлично! При 10V/см ни перегрева, ни закисления или защелачивания. Очень рекомендую! * ubMed will retrieve 2 records. J Virol Methods. 2010 May 13. [Epub ahead of print]Fast short-fragment PCR for rapid and sensitive detection of shrimp viruses. Mrotzek G, Haryanti, Koesharyani I, Tretyakov AN, Sugama K, Saluz HP. Leibniz Institute for Natural Product Research and Infection Biology, Hans Knoell Institute, Beutenbergstrasse 11a, 07745 Jena, Germany; Friedrich-Schiller-University, Jena, Germany. This article describes a fast short-fragment PCR method for the detection of white spot syndrome virus (WSSV), infectious hypodermal and hematopoietic necrosis virus (IHHNV), and monodon baculovirus (MBV). Fast two-temperature (95 degrees C denaturation and 60 degrees C annealing/extension) PCRs were performed in 5-10mul volume samples in miniaturized microplates using a fast Peltier thermal cycler. 40 cycles were completed in 25-30min. Rapid high-resolution agarose gel electrophoresis of 70-150bp PCR fragments was performed in 10min. High sensitivity of PCR product detection (50-100pg) was obtained using ultra sensitive dyes such as GelStar(®) and a gel documentation system equipped with a blue-light transilluminator. This novel method is faster and more sensitive than its TaqMan(®) real-time PCR counterparts. Copyright © 2010. Published by Elsevier B.V. да уж сам себя не похвались как оплеванный ходишь. © источник Электрофорез (от электро- и др.-греч. φορέω — «переношу») —метод разделения макромолекул, различающихся по размеру, молекулярной массе, пространственной конфигурацией, вторичной структурой или электрическому заряду. Впервые было открыто профессорами Московского университета П. И. Страховым и Ф. Ф. Рейссом в 1809 году. Молекулы в буферном растворе обладают электрическим зарядом, величина и знак которого зависят от pH среды. При пропускании электрического тока через раствор в нем формируется направленное электрическое поле, напряженность которого измеряется разностью потенциалов по концам емкости, в которой производится электрофорез. Под действием поля молекулы начинают движение в направлении катода или анода. Скорость движения зависит от величины заряда, размеров и трения окружающей среды. С течением времени смесь разделяется на фракции, состоящие из молекул, движущихся с одинаковой скоростью. В современных экспериментах рабочий канал приборов для электрофореза заполняют гелем, имеющим структуру сетки. В этом случае основное влияние на подвижность молекул и их степень разделения оказывают их линейные размеры. В некоторых случаях может возникнуть ситуация, при которой особо крупные молекулы не проходят через поры геля.
Бромфеноловый синий и ксиленцианол — могут заметно мешать наблюдению фрагментов под UV. Краситель Cresol red совместим с ферментативными реакциями, практически не мешает наблюдению под UV. OrangeG наиболее подвижный краситель, практически всегда находится вне «рабочей зоны». Заметен под UV. Краситель в буфере нужен лишь для того, чтобы образец был легко заметен в лунке и в геле. Самое широкое применение агарозные гели имеют в исследованиях, связанные с разделением нуклиновых кислот. Последние имеют довольно значительные отрицательный заряд, величина которого слабо зависиот от pH раствора, вследствие чего разделение на фракции происходит в основном за счет различия в линейных размеров молекул. В таких экспериментах используют 0.089М Трис-боратный, 0.05 Трсфосфатный и Трис-ацетатный буфер. Стоит отметить, что при обычном электрофорезе в геле можно разделять фрагменты нуклеиновых кислот, размер которых менее 50 тыс п.н. Также часто из эксперимента нужно получить оценку размеров молекул. Для этого используются наборы молекул известной длины. Например, для регистрации продуктов амплификации ДНК применяется электрофорез в агарозном геле в присутствии бромистого эитидия, который образует с фрагментами ДНК устойчивое соединение внедрения, проявляющееся в виде светящихся полос при облучении геля УФ-излучением длиной волны 290-330 нм. источник
Электрофорез в агарозном геле является методом гель — электрофореза , используемого в биохимии , молекулярной биологии , генетики и клинической химии , чтобы отделить смешанную популяцию макромолекул , таких как ДНК или белков в матрице из агарозы , один из двух основных компонентов агара . Белки могут быть разделены с помощью заряда и / или размером ( изоэлектрофокусирование электрофореза в агарозном, по существу , не зависит размер), а также ДНК и РНК — фрагменты длиной. Биомолекулы разделены путем применения электрического поля для перемещения заряженных молекул через матрицу агарозы, и биомолекулы отделены друг от друга размером в матрице агарозном геле. Гель агарозов легко отливать, имеет относительно меньшее количество заряженных групп, и особенно подходит для разделения ДНК диапазона размеров наиболее часто встречающегося в лабораториях, на долю которого приходится популярности ее использование. Разделены ДНК может быть просмотрен с красителем, чаще всего под действием УФ-света, и фрагменты ДНК могут быть извлечены из геля с относительной легкостью. Большинство гели агарозы используются между 0.7-2%, растворенного в подходящем буфере для электрофореза.
Агарозном гель является трехмерный матрицей , составленной из спиральных молекул в агарозных суперспиральных пучках , которые объединяются в трехмерный структуры с каналами и порами , через которые могут проходить биомолекулы. Структура 3-D проводится совместно с водородными связями и , следовательно , может быть разрушена путем нагрева обратно в жидкое состояние. Температура плавления отличается от температуры желатинизации, в зависимости от источников, агарозный гель имеет температуру гелеобразования 35-42 ° C и температуру плавления 85-95 ° С. Низкое плавление и низкие желирующие агарозы , сделанные с помощью химических модификаций, также доступны. Агарозном гель имеет большой размер пор и хорошую прочность геля, что делает его пригодным в качестве антиконвекции среды для электрофореза ДНК и крупных белковых молекул. Размер пор 1% геле была оценена от 100 нм до 200-500 нм, а его прочность геля позволяет гели в виде разбавленных в 0,15% с образованием сляба для гель — электрофореза. Низкая концентрация-гели (0,1-0,2%) , однако являются хрупкими и , следовательно , трудно управлять. Гель агарозы имеет более низкую разрешающую способность , чем полиакриламидный гель для ДНК , но имеет больший диапазон разделения, и поэтому используется для фрагментов ДНК обычно 50-20,000 п.н. в размере. Предел разрешения для стандартного электрофореза в агарозном геле составляет около 750 кб, но разрешение более 6 Мб возможны с электрофорезом геля импульсным поля (PFGE). Он также может быть использован для разделения больших белков, и это является предпочтительным матрица для гель — электрофореза частиц с эффективным радиусом больше 5-10 нм. 0,9% агарозный гель имеет поры достаточно большой для ввода бактериофага Т4 . Агарозном полимер содержит заряженные группы, в частности пирувата и сульфата . Эти отрицательно заряженные группы создают поток воды в направлении, противоположном направлению движения ДНК в процессе , называемом electroendosmosis (РВЗ), и , следовательно , может замедлить движение ДНК и вызывает размывание полос. Более высокие концентрации гели будут иметь более высокий электроосмотический поток. Низкая РВЗ агарозу поэтому обычно предпочтительным для использования в агарозном гель — электрофореза нуклеиновых кислот , но высокая РВЗ агарозы может быть использована для других целей. Ниже , содержание сульфатов низкой РВЗ агарозы, в частности , низкой температурой плавления (LMP) агарозы, также полезно в тех случаях , когда ДНК экстрагировали из геля , которые будут использоваться для дальнейших манипуляций в присутствии загрязняющих сульфаты может влиять на некоторые последующие процедуры, такие , в качестве лигирования и ПЦР . Нулевые РВЗЫ агарозы однако являются нежелательными для некоторых применений , так как они могут быть сделаны путем добавления положительно заряженных групп и такие группы могут повлиять на последующие реакции фермента. Electroendosmosis является причиной агарозном используется в предпочтении к агар как агаропектин компонент в агар содержит значительное количество отрицательно заряженных сульфатов и карбоксильных групп. Удаление агаропектина в агарозе существенно снижает РВЗ, а также уменьшения неспецифической адсорбции биомолекул на гелевой матрице. Тем не менее, для некоторых приложений , таких как электрофорез белков сыворотки, высокая РВЗ может быть желательной, и агаропектин может быть добавлен в геле используется. Ряд факторов может повлиять на миграции нуклеиновых кислот: размерность поры геля (концентрация геля), размер ДНК будучи электрофорез, напряжение используется, ионная сила буфера, а концентрация интеркалирования красителя, такие как бромид этидий если используется во время электрофореза. Более мелкие молекулы движутся быстрее , чем более крупные молекулы в геле, и движется двухцепочечной ДНК со скоростью, обратно пропорциональна логарифму числа пар оснований. Это соотношение , однако ломается при очень больших фрагментах ДНК, и разделение очень больших фрагментов ДНК требует использования электрофореза импульсного геля — поля (PFGE), который применяет переменный ток с двух разных направлений и большие ДНК — фрагменты разделены , как они переориентироваться с изменение тока. Для стандартного электрофореза в агарозном геле, более крупные молекулы разрешаются лучше, используя низкую концентрацию геля в то время как меньшие молекулы отделить лучше при высокой концентрации геля. Высокие концентрации геля, однако, требует больше времени запуска (иногда дней). Движение ДНК может влиять на конформации молекулы ДНК, например, суперспирализовано ДНК , как правило , двигается быстрее , чем расслабленный ДНК , поскольку она плотно спиральная и , следовательно , более компактным. В нормальной подготовке плазмидной ДНК, множественные формы ДНК могут присутствовать. Гель — электрофорез плазмид обычно показывает отрицательно суперспиральную форму как основная полоса, в то время как зазубрины ДНК (открытая круговая форма) и расслаблена закрыт круглая форма появляются как незначительные полосы. Скорость , с которой различными формами перемещения , однако , может изменяться с использованием различных условий электрофореза, а подвижность большой кольцевой ДНК могут быть более сильно , чем влияние линейной ДНК размера пор гель. Этидий бромид, который интеркалирует в кольцевую ДНК может изменить заряд, длину, а также superhelicity молекулы ДНК, поэтому его присутствие в геле при электрофорезе может повлиять на его движение. Например, положительный заряд бромистого этидия может уменьшить движение ДНК на 15%. Электрофорез в агарозном геле может быть использован для решения круговой ДНК с различной топологией суперспирализации. Повреждение ДНК из — за увеличенное поперечное сшивание также приведет к уменьшению электрофоретической миграции ДНК в зависимости от дозы. Скорость миграции ДНК пропорциональна приложенного напряжения, т.е. чем выше напряжение, тем быстрее движется ДНК. Разрешение больших фрагментов ДНК, однако ниже, при высоком напряжении. Подвижность ДНК может также изменяться в нестационарном поле — в поле, которое периодически обращено, подвижность ДНК определенного размера может значительно снижаться при определенной частоте езды на велосипеде. Это явление может привести к инверсии зоны в поле электрофореза гель инверсии (ФИГЕ), в результате чего более крупные фрагменты ДНК движутся быстрее, чем более мелкие.
Отрицательный заряд его остова ДНК перемещает в сторону положительно заряженного анода при электрофорезе. Тем не менее, миграция молекул ДНК в растворе, в отсутствии гелевой матрицы, не зависит от молекулярной массы при электрофорезе. Гелевая матрица, следовательно , отвечает за разделение ДНК по размерам в процессе электрофореза, а также ряд моделей существует , чтобы объяснить механизм разделения биомолекул в гелевой матрице. Широко принято одна модель Ogston , которая рассматривает матрицу полимера в виде сита. Шаровые белки или случайная катушка ДНК перемещается через взаимосвязанные поры, и движение больших молекул, более вероятно, будет затруднено и замедлены столкновениями с гелевой матрицей, и , следовательно , молекулы различных размеров могут быть отделенны в этом процессе просеивания , Модель Ogston однако ломается при больших молекул в результате чего поры значительно меньше , чем размер молекулы. Для молекул ДНК размера более 1 кб, А рептации модель (или его варианта) наиболее часто используется. Эта модель предполагает , что ДНК может сканировать в моде «змеиной» (отсюда «рептации») через пору в виде удлиненной молекулы. Необъективное модель рептаций применяется при более высокой напряженности электрического поля, в результате чего передний конец молекулы становится сильно предвзятым в прямом направлении и тянет остальную часть молекулы вместе. В режиме реального времени флуоресцентной микроскопии окрашенных молекул, однако, показали более тонкие динамики во время электрофореза с ДНК показывает значительную эластичность , как она попеременно растяжения в направлении приложенного поля , а затем договаривающуюся в шарик, или становится замкнув в U-образную форму когда он попадает на полимерных волокон. Детали электрофорезом на агарозном геле эксперимента могут варьироваться в зависимости от методов, но большинство следовать общей процедуре. источник Для визуализации результатов операций, проводимых с ДНК,таккак выделение, рестрикция,полимеразная цепная реакция(ПЦР),молекулярное клонирование, наиболее частоиспользуютэлектрофорез. Электрофорез — метод разделения макромолекул (в том числе Для проведения горизонтального электрофореза используют пластину агарозного геля необходимой концентрации с добавлением специального красителя ДНК, например бромида этидия. На скорость движения ДНК в геле в процессе электрофореза влияют несколько факторов. Концентрация агарозы в геле.Агарозный гель — пористая струк- Существует зависимость длины разделяемых фрагментов ДНК Концен- 0,3 0,5 0,6 0,7 0,8 0,9 1,0 1,2 1,5 2,0 Длина 5-60 1-30 1-20 0,8-12 0,6-10 0,5-8 0,5-7 0,4-6 0,2-3 0,1-2 Заряд молекулы.Поскольку каждый из нуклеотидов молекулы Напряженность электрического поля.На скорость движения заряженных молекул ДНК в геле влияет напряженность электрического поля, определяемая напряжением постоянного электрического поля, подаваемого на электроды. Данные, приведенные на Линейные молекулы ДНК одного размера движутся в геле с • суперспирализованную (нативная молекула, стабилизированная белками); •линейную (расщепленная рестриктазой кольцевая молекула). Кольцевая молекула ДНК При постановке электрофореза можно определить размер (молекулярную массу) только линейной ДНК. Для этого в одну из лунок геля наносят стандарт, в качестве которого используют специальные маркеры молекулярной массы (смесь фрагментов ДНК с Для контроля скорости движения ДНК в геле, а также для определения времени окончания процесса электрофореза применяют краску-лидер (специальный краситель, например бромфеноловый синий), которая перемещается в геле, немного опережая макромолекулы ДНК, двигающиеся в процессе электрофореза. Для визуализации результатов электрофореза используют краситель бромид этидия, который вносят в процессе приготовления геля. Данное вещество встраивается (интеркалирует) в молекулы ДНК плоскими ароматическими группами. После окончания Внимание! Используемый в качестве красителя бромид этидия является мутагенным веществом. При работе с ним необходимо использовать резиновые или латексные перчатки. Методы вертикального и горизонтального электрофореза Кроме того,акриламид является токсичным веществом. Приготовить ПААГ Цель работы. Ознакомиться с методом горизонтального электрофореза ДНК в агарозном геле. Оборудование и материалы.1. Прибор для горизонтального электрофореза. Ходработы.Приготовлениерабочегобуферногораствора для электрофореза. Содержимое пакета «Буфер для электрофореза» полностью переносят в мерную колбу, раство-ряют в 600—800 мл дистиллированной воды (для более быстрого Подготовка прибора для электрофореза к работе. Пользуясь встроенными уровнями и винтовыми ножками, высоту аппликатора выставляют таким образом, чтобы расстояние Приготовление агарозного геля. Навеску агарозы, Проведение электрофореза. В лунки застывшего агарозного геля осторожно вносят по 3 мкл раствора исследуемых образцов ДНК. В соседнюю лунку геля вносят 3 мкл маркера молекулярных масс фрагментов ДНК. В одну или Внимание! Во избежание повреждения сетчатки глаз ультрафиолетовым излучением наблюдать зоны ДНК следует только через за-щитное стекло из комплекта трансиллюминатора или защитные (стеклянные) очки. Полученные результаты регистрируют визуально или с использованием гель-документирующей видеосистемы,пользуясь инструкцией к ней. Контрольные вопросы.1. Какой принцип лежит в основе метода электрофоре- Задания.1. Поместить схему или фотографию электрофореграммы в рабочий журнал, пронумеровать дорожки и сделать подписи к ним. 2. Определить и подписать размеры фрагментов ДНК-маркера (согласно описанию к нему). 3. По электрофореграмме определить электрофоретическую подвижность и рассчитать примерную молекулярную массу исследуемой плазмидной ДНК, используя маркеры молекулярных масс. 4. Определить, какой из ДНК нижеперечисленных плазмид соответствует исследуемая ДНК, если ДНК плазмиды РСЕМ-2Гимеет размер 3000 п.н.ДНК плазмиды рВК322 — 4361,а ДНК плазмидыр РСУ002—8560 п.н. источник Метод электрофореза в геле использует разницу в размере и заряде различных молекул в образце. Образец ДНК или белка, подлежащий разделению, погружают в пористый гель, помещенный в ионную буферную среду. При приложении электрического поля каждая молекула, имеющая разный размер и заряд, будет проходить через гель с разной скоростью. Пористый гель, используемый в этой технике, действует как молекулярное сито, которое отделяет большие молекулы от более мелких. Меньшие молекулы движутся быстрее по гелю, а более крупные медленнее. Подвижность частиц также определется их индивидуальным электрическим зарядом. Два противоположно заряженных электрода, которые являются частью системы, тянут молекулы к себе на основе их заряда.
Образцы, которые необходимо проанализировать, затем загружают в маленькие лунки в геле с помощью пипетки. По завершении загрузки применяется электрический ток 50-150 В. Теперь заряженные молекулы, присутствующие в образце, начинают мигрировать через гель к электродам. Отрицательно заряженные молекулы движутся к положительному электроду, а положительно заряженные молекулы мигрируют к отрицательному электроду. Скорость, с которой каждая молекула перемещается через гель, называется ее электрофоретической подвижностью и определяется главным образом ее чистым зарядом и размером. Сильно заряженные молекулы движутся быстрее, чем слабо заряженные. Меньшие молекулы работают быстрее, оставляя более крупные. Таким образом, сильный заряд и малый размер увеличивают электрофоретическую подвижность молекулы, а слабый заряд и большие размеры уменьшают подвижность молекулы. Когда все молекулы в образце имеют одинаковый размер, разделение будет основываться исключительно на их размере.
|
 Электрофорез в агарозном геле в различных модификациях широко применяется для разделения молекул нуклеиновых кислот, белков и других макромолекул в биологии и медицине. На водяной бане или в лабораторной печи плавят смесь агарозы, буфера и воды. Затем ее охлаждают до 50-60 °C и заливают в форму. Лунки для нанесения делаются при помощи гребенки. Исследуемый образец наносят в лунку при помощи дозатора. Когда краситель, помещаемый в лунки в начале эксперимента, достигает конца геля, электрофорез останавливают. Затем гель окрашивают красителем, который связывается с исследуемыми молекулами. Интенсивность окраски полос красителя дает представление о концентрации молекул в образце. Кроме концентрации, метод электрофореза позволяет определить относительную молекулярную массу исследуемых молекул, для этого в крайнюю лунку помещают набор маркеров молекулярной массы, который должен полностью покрывать диапазон молекулярных масс исследуемой системы.
Электрофорез в агарозном геле в различных модификациях широко применяется для разделения молекул нуклеиновых кислот, белков и других макромолекул в биологии и медицине. На водяной бане или в лабораторной печи плавят смесь агарозы, буфера и воды. Затем ее охлаждают до 50-60 °C и заливают в форму. Лунки для нанесения делаются при помощи гребенки. Исследуемый образец наносят в лунку при помощи дозатора. Когда краситель, помещаемый в лунки в начале эксперимента, достигает конца геля, электрофорез останавливают. Затем гель окрашивают красителем, который связывается с исследуемыми молекулами. Интенсивность окраски полос красителя дает представление о концентрации молекул в образце. Кроме концентрации, метод электрофореза позволяет определить относительную молекулярную массу исследуемых молекул, для этого в крайнюю лунку помещают набор маркеров молекулярной массы, который должен полностью покрывать диапазон молекулярных масс исследуемой системы.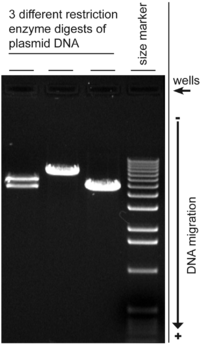

 Гель, используемый в геле-электрофорезе, обычно изготавливают из материала, называемого агарозой, который представляет собой желатиновое вещество, экстрагированное из водорослей. Этот пористый гель можно использовать для отделения макромолекул разных размеров. Гель погружают в раствор солевого буфера в камеру электрофореза. Трис-борат-ЭДТА (ТВЭ) обычно используется в качестве буфера. Его основная функция — контролировать pH системы. Камера имеет два электрода — один положительный и другой отрицательный — на двух концах.
Гель, используемый в геле-электрофорезе, обычно изготавливают из материала, называемого агарозой, который представляет собой желатиновое вещество, экстрагированное из водорослей. Этот пористый гель можно использовать для отделения макромолекул разных размеров. Гель погружают в раствор солевого буфера в камеру электрофореза. Трис-борат-ЭДТА (ТВЭ) обычно используется в качестве буфера. Его основная функция — контролировать pH системы. Камера имеет два электрода — один положительный и другой отрицательный — на двух концах. источник
источник